

◆視覚が生じる前の網膜活動が緻密な神経回路網を設計する
◆中年太りを引き起こす神経細胞のかたちの変化
◆Ca2+やcAMP を感知する蛍光タンパク質を開発
◆抗うつ作用に重要な脳領域を発見
◆報酬とリスクの意思決定バランスを光で調節 ―精神神経疾患の病態解明に期待―
◆神経回路リモデリングにおいて特定のニューロン構造を選択的に除去するメカニズム
◆糖の摂食により痛覚応答が抑えられる仕組みを解明 ――個体の栄養状態に応じた末梢痛覚のチューニング――
◆リズム知覚における小脳と大脳基底核の機能の違い
◆眠りの量と眠気を制御するリン酸化経路
◆うつ症状を生じる脳内メカニズムとその制御法
◆闇夜に揺れる柳から、なぜ逃げ出してしまうのか? ― 恐怖と θ 活動の話
◆統合失調症を引き起こす巨大なシナプス
◆同じ失敗を繰り返さないために必要な脳内メカニズムの解明
◆大人の脳で作られた神経細胞のシナプスの数を調節する仕組みを発見
◆マウス脳における複雑な視覚神経ネットワークの形成過程を解明
◆挑発を受けると攻撃的になる脳内の仕組み
◆体の「痛い」を脳から治す ―痛みに関わる神経回路を標的とした疼痛の新たな治療戦略―
◆体温の中枢調節の基本原理
◆言葉が示す内容と記された色の矛盾を乗り越えるための脳のしくみ
◆ラットも音楽のビートに合わせて身体を動かすことを発見
◆運動指令信号と感覚信号が統合されて運動が作り出される過程を発見
◆シナプス貪食を介した神経回路の最適化
◆シナプスの個性を決める分子群の微細な集積構造 -脳標本とともに『膨らむ』神経科学研究の夢-
◆記憶をアップデートする仕組みを解明
◆シナプス集積によって情報統合のダイナミックレンジを広げる仕組み
◆オスマウスのフェロモンがオス同士の争いを引き起こす神経メカニズム
◆父親の子育てを支える神経回路の変化
◆リズムに合わせて動くための小脳による予測的な運動制御メカニズム
◆レム睡眠の開始機構を解明 ~睡眠周期の生成に関するドーパミンと扁桃体の新たな役割の発見~
◆記憶の獲得によって脳領域横断的に情報を統合する ネットワークが形成される
◆発症1週間後の脳梗塞マウスで治療効果を発揮する蛋白質徐放性ゲル化ペプチドの開発
◆温度を感じる分子がストレス応答に関わるしくみ
◆光で記憶を消去する―記憶に睡眠が必要な理由を解明―
◆繰り返し見た画像であれば見にくくなっても知覚できる脳の仕組み
◆水と油の関係でわかった記憶形成の分子機構
◆女性ホルモンが「かゆみ」の感受性を変えるしくみ
◆脳梗塞による細胞死を抑える分子メカニズム
◆X線を使った脳神経操作法の開発
◆「視覚的な動き」はまず網膜の神経軸索終末で検出される
◆視覚野は外界情報だけでなく、動物の内的な状態も表現する
◆脳はハブ細胞が存在するエコなシステムであった
◆「用意,ドン」フライングせずに行動を準備する脳内メカニズム
◆相互に抑制する扁桃体抑制性神経核による恐怖状態の協調制御
◆脳が完成するまでに「生き残る」回路と「刈り込まれる」回路との違いを解明
◆未来の行動に先立って成功確率を予測する仕組みを解明
◆空間認識を支える脳情報の流れを解明
◆生まれたての神経細胞が旅立つ最初期ステップを解明~脳室面に付着した神経細胞の足をDSCAMタンパク質が切り剥がす~
◆状況に応じて物の価値判断を変化させる脳の仕組みを解明
◆神経活動を操作する新技術『化学遺伝学』に、飛躍的に性能が向上した薬剤が登場
◆脳内のグリア細胞―神経細胞間コミュニケーションを解き明かす 空間的分子探索技術の創出
◆母性ホルモン・オキシトシンがオスの性機能を促進させる新たな局所神経機構を解明
◆途切れた神経回路を再びつなぐ人工シナプスコネクターを開発 ~シナプス異常による精神・神経疾患の治療に新しい道~
◆冬眠様状態を誘導する神経経路の発見
◆謎の脳領域「前障」は睡眠中の脳活動を制御する
◆塩のおいしさを生み出す細胞とその仕組み
◆心と身体をつなぐ脳の神経回路メカニズム
◆適切な意思決定を可能にする神経回路を同定
◆レム睡眠中の記憶忘却を誘導する視床下部MCH神経
◆視床下部神経回路の周期的なリモデリングによる雌性行動の制御
◆世界最高性能 Ca2+センサー 『XCaMP』開発による脳情報動態の精密解読
◆脳内神経回路再編をもたらすシナプス形成因子の新しい分泌様式を解明 -神経活動に応じたシナプスの「スクラップ&ビルド」-
◆神経活動の時間的なパターンに基づいた神経ネットワーク形成
◆「根気」は海馬とセロトニンが制御する
◆意思決定の個体差を生む脳内機構 -外的撹乱に対する神経活動の応答性の違いが行動選択の個性を決める-
脳内神経回路再編をもたらすシナプス形成因子の新しい分泌様式を解明
-神経活動に応じたシナプスの「スクラップ&ビルド」-
所属 聖マリアンナ医科大学
職名 講師 氏名 井端啓二
私たちの脳内では、約1000億個の神経細胞が互いに結合し、1000兆個ものシナプスを形成することで、記憶や学習に必須な神経回路を構築します。近年、神経細胞の結び目であるシナプスは、経験や学習によって生涯にわたって絶えず改変されることが分かってきましたが、その詳細な仕組みについては不明な点が多く残されています。
運動学習やその記憶を担う小脳には、顆粒細胞やプルキンエ細胞と呼ばれる神経細胞が存在し、多くの顆粒細胞が軸索(平行線維)を介してプルキンエ細胞に投射することでシナプス(平行線維シナプス)を形成しています(図A)。これまで私たちの研究室では、顆粒細胞内で産生されるCbln1と呼ばれるタンパク質が細胞外に分泌され、シナプス前部と後部にそれぞれ発現する受容体分子と三者複合体(Nrx-Cbln1-GluD2複合体)を構築することで平行線維シナプス形成を促すことを明らかにしました(Science, ’16, ’10)。しかし、このCbln1がどのような経路を介して顆粒細胞から分泌されるのかは不明のままでした。この疑問を解くために、私たちはまず顆粒細胞内におけるCbln1の局在様式を調べました。すると、Cbln1は平行線維内に存在する小胞に局在することが分かりました。平行線維内には神経伝達物質を含むシナプス小胞や、神経栄養因子などを含む有芯小胞が存在し、両者ともに神経活動依存的に分泌されることが知られています。そこで、Cbln1の分泌様式を調べたところ、予想通り、神経活動依存的に軸索から分泌されることが分かりました(図B)。興味深いことに、Cbln1が局在する小胞を詳しく調べると、その小胞内には、神経伝達物質や神経栄養因子は存在せず、タンパク質分解酵素であるカテプシンB(CatB)が局在し、神経活動に伴ってCbln1とCatBが小胞内から一緒に分泌される様子が観察されました(図B)。CatBは細胞内の不要なタンパク質を分解する場であるライソソームに局在する分子です。そこで最後に、ライソソームから細胞外への分泌過程を人為的に阻害すると、面白いことに、神経活動に応じたCbln1とCatBの分泌が抑えられるとともに、マウス小脳においてシナプスのもととなる部分の数が有意に減少しました。つまり、Cbln1はライソソームに局在し、神経細胞が活性化するとタンパク質分解酵素と共に細胞外に分泌され、何らかの細胞外基質を破壊し(スクラップ)、そして創造(ビルド)することでシナプス形成を促進していたのです(図C)。
この神経活動に応じたシナプスのスクラップ&ビルド現象は、生涯にわたって観察される神経回路再編を引き起こす重要な過程であると考えられ、将来、本成果が、記憶・学習の分子的理解だけでなく、シナプス不全を伴う様々な精神神経疾患に対する治療法開発につながるものと期待されます。
Activity-dependent secretion of synaptic organizer Cbln1 from lysosomes in granule cell axons.
Keiji Ibata, Maya Kono, Sakae Narumi, Junko Motohashi, Wataru Kakegawa, Kazuhisa Kohda, Michisuke Yuzaki. (2019) Neuron 102(6):1184-1198.
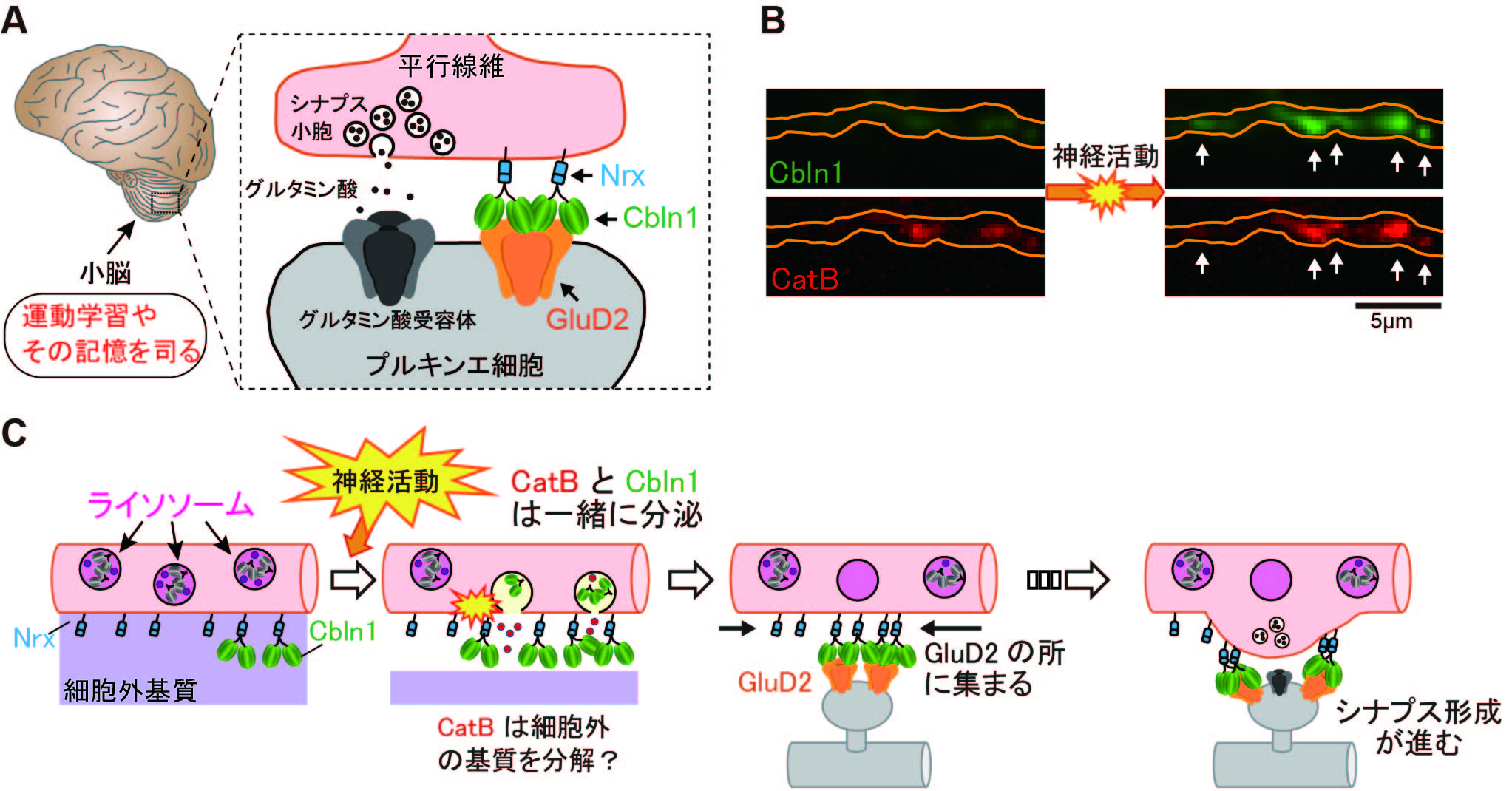
<図の説明>
(A) 小脳平行線維シナプスをつなぐ三者複合体構造
小脳顆粒細胞の平行線維から分泌されるCbln1が、平行線維上のニューレキシン(Nrx)およびプルキンエ細胞上のδ2型グルタミン酸受容体(GluD2)と結合することによって、平行線維シナプスを形成する。
(B) 神経活動によって平行線維からCbln1とCatBが分泌される
細胞外に分泌されると明るくなる蛍光タンパク質でラベルしたCbln1(緑、上段)とCatB(赤、下段)を発現する平行線維。神経活動を活性化すると右側矢印のように蛍光が明るくなった。
(C) 本研究の概要
(左から順に)
・シナプス形成を誘導するCbln1は、平行線維内のライソソームに存在する。
・神経活動によってライソソームが細胞膜と融合し、CatBなどのライソソーム酵素とともにCbln1が細胞外に放出される。
・放出されたCbln1は平行線維上のNrxに結合して拡散しGluD2を発現するプルキンエ細胞と接触した場所に集まり、Nrx-Cbln1-GluD2複合体が形成される。
・複合体が形成された領域でシナプスの形成が進み、成熟シナプスが出来上がる。
<研究者の声>
本研究は、慶應義塾大学の生理学柚﨑研究室に所属した時からスタートして、8年ほど掛けて仕上がった論文です。研究環境がとても恵まれていたので、任期や業績欄等のことは忘れ、良い研究を行うことだけを考えて実験を進めました。足りないデータはまだまだあったと思いますが、無事に受理され安堵しています。長い間ご指導くださった柚﨑先生やサポートいただいた方々に深く感謝いたします。
<略歴>
2001年、東京大学大学院医学研究科(御子柴研究室)にて博士号取得
2001年、米国、ブランダイス大学(Turrigiano研究室)ポスドク
2006年、理化学研究所脳科学総合研究センター 細胞機能探索技術開発チーム(宮脇研究室)研究員
2010年、慶應義塾大学医学部生理学(柚﨑研究室)特別研究助教、2012年より生理学助教
2017年、聖マリアンナ医科大学生理学(幸田研究室)助教、2019年より講師




HOME | 一般社団法人 日本神経科学学会
- 学会機関紙
- Neuroscience Research
- Articles in Press
- Latest Issue
- Back Issues
- Submit Your Paper
- About the Journal
- 電子版の購読について
(会員の方へ) - お知らせ
- 公募情報 助成・受賞
- 研究員・教職員募集
- 大学院生募集・説明会
- 研究助成・渡航助成募集
- 受賞候補者募集
- キャリアパスに応じた賞・助成
- 海外の学会への旅費支援
- 学会推薦による賞・助成
- 成茂神経科学研究助成基金
- イベント・研究会情報
- イベント・研究会
- 一般の方向けイベント
- 神経科学の発展のために
- 倫理問題などに関する指針
- 動物実験に関する指針
- MRI検査の指針
- ランチョン大討論会
- ダイバーシティの取り組みについて
- 会員ページ
- Neuroscience Research (NSR) 電子版閲覧
- 抄録検索システム [IDパスワードが必要]
- 会員ログイン
- 過去の神経科学ニュース